
{kind=link}
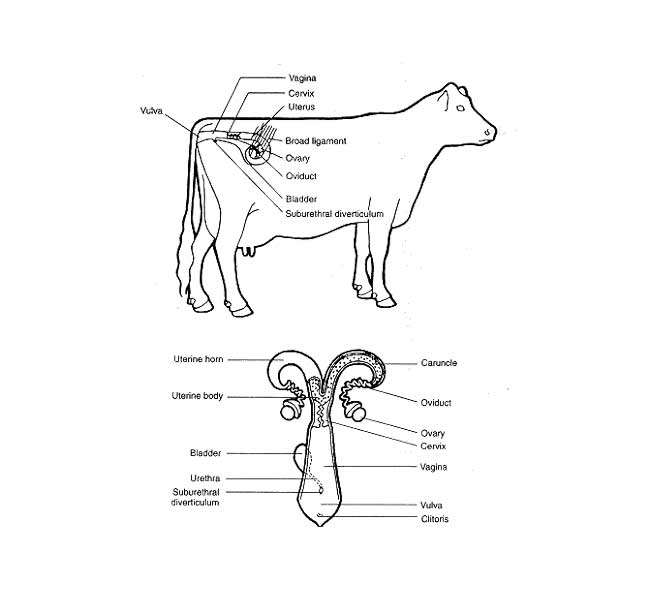
Döl verimin fizyolojisi. Üreme fizyolojisinin temellerinin iyi bilinmesi özellikle ineklerdeki infertilite sorununun teşhisi ve sağıtımı açısından çok önemlidir. Başta follikül gelişmesi olmak üzere gametlerin taşınması ve birbirleri ile kaynaşması (fertilizasyon) olguları hormonların denetimi altındaki son derece karmaşık mekanizmalar sonucu oluşur. O yüzden, follikül’lerin gelişmesini yada gametlerin taşınmasını engelleyici fizyolojik olmayan her etken infertilite’ye neden olarak ineklerde önemli ekonomik kayıplara yol açmaktadır.
Folikülogenezis
Yeni doğan bir dişi buzağının ovarium’larında 75.000 ile 150.000 arasında değişen sayıda ilkel follikül ve oogonium rezervi bulunur. Doğumdan kısa süre önce follikül ve oosit gelişmesi eş zamanlı olarak başlar. Bu sırada oogonium’lar mitoz bölünme ile çoğalarak primer follikül’ler içinde primer oosit’ler şekillenir. Ergenlikten sonra primer oosit’ler meyoz ile bölünerek kromozom sayısının yarıya indiği sekunder oosit’ler ve sekunder (seçilmiş, preantral) follikül’ler, onların da mitoz ile bölünmesi sonucu tersiyer oosit’ler ve tersiyer (dominant, antral) follikül’ler meydana gelir.
Follikül gelişmesi ergenlikten sonra dişi üreme kanalında genetik, patolojik, enfeksiyon sorunu bulunmayan ; güneş ışığı ve oksijeni yoğun açık sistem ahırlarda barındırılan; vitamin, mineral, iz elementten zengin enerjili yemlerle beslenen ineklerde gebelik, involusyon, diöstrus evreleri de dahil olmak üzere tüm cinsel yaşam boyunca kesintisiz olarak devam eder. Yılda tek yavru doğuran ineklerde her kızgınlık siklusu’nda gelişen follikül’lerden sadece biri ovum taşıyan Graff follikülü haline dönüşür, diğerleri gerileyerek yok olur.
Ergenliğe ulaşan bir dişi buzağıda follikül gelişmesi ile hormon salgılanması eş zamanlı olarak başlar. Hormon salgılanmasını uyaran etkenler çok ve çeşitlidir. Bunlar arasında en başta hayvanın yaşını ve canlı ağırlığını sayabiliriz. Kültür ırkı inekler genelde 12 aylıkken ergenliğe ulaşırlar. Ancak yaş ergenliğe ulaşmada tek etken de değildir. İneğin ergin yaş ağırlığının %40-50’ sine ulaşması ve vücut kondisyonunun yeterli olması gerekir. Yeterli canlı ağırlık ve uygun vücut kondisyonuna sahip ineklerde üreme organlarının gelişmesi de zamanında olur. Öte yandan çevre ısısı, gün ışığı, boğa kokusu (pheromen) gibi ineğin duyu organları ile duyumsayabileceği etkenler de hormon salgılanmasının dolayısıyla follikül gelişiminin başlamasına katkıda bulunur. Örneğin güneş ışığı retina ve nervus opticus yoluyla beyindeki hipotalamus’a gelerek buradaki nöronlardan inekteki tüm üreme işlevlerini başlatan ve denetleyen Gonadları Uyarıcı Hormon (Gonadotropik Releasing Hormon-Gn-RH) salgılatır.
Yukarıda sayılanlara ek olarak Gn-RH salgısını uyaran önemli bir etken de Insulin Like Growth Factor (İnsulin Benzeri Büyüme Faktörü , IGF-1)’ dür. Özellikle düşük enerjili yemlerle beslenen ineklerde IGF-1 dolayısıyla da Gn-RH salgılanamamakta, bu da ileride ayrıntılarıyla üzerinde duracağımız gibi kimi infertilite olgularına neden olmaktadır.
Gn-RH hipotalamus’tan periyodik dalgalar halinde salgılanır. Her Gn-RH dalgası FSH ve LH’nın da hipofiz’den dalgalı biçimde salgılanmasına yol açar. Gn-RH özel reseptörlere bağlanarak kan yoluyla hipofiz ön lobuna gider ve oradaki gonadotrop hücreleri uyararak FSH ve LH’nın salgılanmasını doğurur. FSH, follikül’leri dominant (preantral) follikül aşamasına kadar geliştirir. LH ise dominant follikül’lerin olgunlaşmasını, antrum şekillenmesini ve ovulasyonu doğurur. FSH follikül’deki granulosa hücrelerini uyararak östradiol’un, LH da teka hücrelerini uyararak testesteron’un salgılanmasına neden olur. Östradiol ve testesteron arasındaki denge follikül’lerin fizyolojik bütünlüğünü ve canlılığını sağlar.
Düve ve inekler her siklusta 2, 3, hatta 4 follikül gelişim dalgasına sahiptir. Her bir follikül dalgası 7-11 günlük bir süreyi kapsar. İki follikül gelişim dalgasına sahip ineklerde birinci dalga siklusun 2. gününde (1.-2. günler arası) , ikinci dalga ise 11.günde gelişmeye başlar ve hemen sonra ovulasyon şekillenir. Üç follikül gelişim dalgasına sahip ineklerde birinci dalga 2. günde gelişir, 8.-12. günlerde statik hale gelir ve 12.-16. günlerde follikül gerileyerek yok olur. İkinci dalga 10.-11. günlerde başlar ve 17.günde sona erer. Üçüncü dalga ise 16.günde başlar ve ovulasyonla sonlanır.
Gn-RH ‘nın salgılanmasına neden olan ve baştan beri sıralanan dış etkenler yanında denetlenmesini sağlayan iç etkenler de vardır. Progesteron, östrogen , FSH ve LH gibi hormonların karşılıklı olumlu yada olumsuz etkileşimleri sonucu Gn-RH salgılanması dolayısıyla da follikül gelişmesi denetlenmektedir.
Ovum Olgunlaşması
Memeli oositleri iki aşamada oluşmaktadır.
1) Gelişme Aşaması: Bu aşamada follikül rezervden ayrılır, gelişmeye ve oosit’le birlikte büyümeye başlar, antrum şekillendiğinde gelişme tamamlanmış olur,
2) Nukleer ve Sitoplazmik Hazırlanma Aşaması: Oosit’in fertilizasyona hazırlanma evresidir. Burada iki çeşit hücre bölünmesi ayırt edilir. Birincisi vücudun soma hücrelerinde daha çok görülen mitoz bölünmesi, diğeri de sadece gametlere özgü olan meyoz yada redüksiyon bölünmesidir. Yukarıda belirtildiği gibi folliküller’de oogonium’lar mitozla bölünmek suretiyle primer oosit’lere, primer oosit’ler meyozla bölünmek suretiyle sekunder oosit’lere, sekunder oosit’ler de tekrar mitozla bölünmek suretiyle tersiyer oosit’lere dönüşürler.
Döl Verimin Fizyolojisi Ovulasyon
Ovulasyonun oluşum evrelerinin ve özellikle de hormonal mekanizmasının iyi bilinmesi ineklerde infertilite olgularının teşhisini ve sağıtımını kolaylaştıracak önemli bir olgudur.
Ovulasyon, olgunlaşmış daha doğrusu patlamaya hazır hale gelmiş Graff follikülü’nün çatlayarak cumulus ooforicus içerisinde bulunan, nukleer ve sitoplazmik olgunlaşmasını tamamlamış ovum’un etrafındaki granulosa hücrelerinden oluşan corona radiata katı ile birlikte oviduct’un fimbria ovarica bölümü tarafından alınarak fertilizasyon bölgesine nakledilmesi olayıdır.
Döl Verimin Fizyolojisi olarak follikül ovulasyon sırasında başlıca üç büyük değişikliğe uğrar.
- oosit’in nukleus ve sitoplazmasının olgunlaşması,
- cumulus ooforicus hücrelerinin granulosa hücre katı boyunca adacık oluşturacak biçimde ayrılması,
- dış folliküler tabakanın incelmesi.
Ovulasyonun oluşması için kimi faktörlerin etkili olması gerekmektedir.
Bu faktörleri şöylece sıralayabiliriz.
1) graff follikülü’nü dolduran sıvının çoğalarak iç basıncın artması,
2) ovarium’daki damarların aşırı kanla dolması sonucu follikül çevresindeki kapillar basıncın artması,
3) biyokimyasal ve farmakolojik faktörler,
4) musküler ve vasküler faktörler,
5) endokrinolojik faktörler,
6) enzimatik faktörler.
Döl Verimin Fizyolojisi İneklerde ovulasyon diğer memeli türlerinden farklı olarak ovarium’un tüm yüzeyi boyunca ve östrusun bitiminden sonraki bir dönemde kendiliğinden oluşur. Luteal fazda yani corpus luteum’un egemen olduğu metöstrus ve diöstrus evrelerinde kanda düzeyi giderek artan progesteron hipotalamus üzerine baskı yaparak Gn-RH dolayısıyla da FSH- LH’ nın salgılanmasını engeller ve follikül gelişmesinin Graff follikülü’ne değin uzamasına engel olur. Ovulasyonu başlatan asıl etken kızgınlık siklusu’nun 16.gününde uterus endometriumu’ndan salgılanan prostaglandin hormonunun etkisiyle corpus luteumun erimesi ve böylece başlayan siklusun proöstrus ve östrus evrelerinde kanda aniden düzeyi düşen progesteron’un Gn-RH üzerindeki baskıyı (negatif feed-back) ortadan kaldırmasıdır. Gn-RH etkisiyle hızla yükselen FSH dalgaları preantral (sekunder) follikülleri alıp, önce antral (tersiyer) sonra da Graff follikülü haline dönüştürür ve sıklığı artan LH dalgaları öteki etkenlerle birlikte ovulasyonu oluşturur.
LH dalgalarının etkisiyle follikül’ün fibroblast hücreleri prostaglandin’leri ve progesteron’u salgılar. Prostaglandin’ler ovarium’lardaki düz kas kontraksiyonlarını , progesteron da teka hücrelerini aktive ederek proteolitik enzimlerin ve kollagenaz’ın salgılanmasını uyarır. Proteolitik enzimler ve kollagenaz nekro dejeneratif olaylar sonucu follikül hücrelerinin dejenerasyonuna neden olarak basal laminayı inceltir. Yırtılmaya neden olan ise prostaglandin’lerin etkisi ile güçlenen ovarium düz kaslarının kontraksiyonudur.
Fertilizasyon
Spermatozon’un ovum’a girmesi ve onunla kaynaşması olarak tanımlanan fertilizasyon (fekondasyon,döllenme), gametlerin oviduct’un ampulla-isthmus bölgesine taşınması, spermatozoon kapasitasyonu ve akrozom reaksiyonu gibi ön gerekleri içeren karmaşık bir olaylar bütünüdür. Fertilizasyona geçmeden önce bu konular üzerinde kısaca durulacaktır.
Gametlerin Taşınımı
Tohumlama sonrası spermatozoon’lar bir yönde güçlü hareketleri ve düz kasların caudo-cranial kontraksiyonu sayesinde üreme kanalında hızla ilerlerler. Bu esnada bir yandan kapasite olurken bir yandan da bakteri toksinleri, ortam pH’sı, anti-sperm agglutinin’leri gibi kimi engelleyici etmenlerle savaşmak zorunda kalırlar. Spermatozoon’lar özellikle cervix uteri’den geçerken büyük kayıplara uğrarlar. Böylece sayıları iyice azalan spermatozoon’lar cornu uteri ile oviduckt’un birleştiği uterus-oviduct bağlantı bölgesini de geçip isthmus’a geldiklerinde kapasitasyon sürecini tamamlarlar ve ampulla-istmus sınırına gelirler. Ovum, fertilizasyon yeri olarak ta tanımlanan bu bölgeye ovulasyondan sonra içinde bulunduğu follikül sıvısının itmesi, cilium’lu epitellerin hareketleri ve infundibulum’un düz kaslarının kontraksiyonu yardımıyla yuvarlanarak ulaşır ve eğer spermatozoon henüz gelmemişse beklemez yoluna devam eder. Ancak, ovum fertilizasyon bölgesini geçip isthmus’a geldiğinde ortamdaki sıvılarla kaplanır ve döllenme yeteneğini yitirir.
Spermatozoon Kapasitasyonu
Boğa spermatozoon’ları testis’ten çıkıp caput epididimis’e gelinceye kadar bağımsız hareket etme ve ovum’la ilişkiye girme yeteneğinde değillerdir. Bu iki özelliği epididimis kanalından geçişleri sırasında kazanırlar ve bu olgu spermatozoon’ların olgunlaşma safhası olarak tanımlanır. Epididimis’teki olgunlaşma safhasında spermatozoon’un nukleus’unda, akrozom’unda ve plazma membranı’nda moleküler değişiklikler oluşur. Ejekülasyon sırasında seminal plazma içinde bulunan glikoprotein yapıdaki makromoleküller plazma mebranı’ndan dış ortama uzanan karbonhidrat zincirleri ile çapraz bağlantı yaparak membranın stabilizasyonuna ve spermatozoon’un dölleme gücünü kaybetmesine neden olurlar.
Ejekülasyondan sonra spermatozoon’ların ovum’la ilişkiye girebilmeleri yani yeniden dölleme gücü kazanabilmeleri için dişi üreme kanalında kapasitasyon süreci geçirmeleri gerekir. Kapasitasyon genel olarak spermatozoon’ların ovum örtülerini geçebilmeleri için gerekli akrozom reaksiyonuna yol açacak fizyolojik-biyokimyasal değişimlerin bütünü olarak tanımlanabilir. Kapasitasyonun oluşumunda asıl önemli unsur bireyin hormon durumudur. Siklusun östrogen’in egemen olduğu proöstrus ve östrus evrelerinde kapasitasyon uyarılırken , progesteron’un egemen olduğu metöstrus ve diöstrus evrelerinde ve yalancı gebelikte engellenir.
Kapasitasyonun moleküler mekanizması henüz tam olarak anlaşılamamış ise de, ejekülasyon sırasında plazma membranına bağlanmış olan glikoprotein yapıdaki makromoleküllerin spermatozon yüzeyinden uzaklaştırılması olarak düşünülmektedir. Kapasitasyon sırasında spermatozoon’ların ovum örtülerini geçebilmeleri için gerekli olan akrozom enzimlerinin inhibitörleri ve corona hücrelerini eritici enzimi (corona penetrating enzyme-CPE) inhibe eden bir madde spermatozoon yüzeyinden uzaklaştırılır. İneklerde kapasitasyon tabii çiftleşmede cerviks ve uterus’ta, suni tohumlamada uterus’ta başlar .
Akrozom Reaksiyonu
Akrozom, spermatogenezis sırasında golgi kompleksi’nden köken alarak gelişen ve nukleus’un yarısını örten takke benzeri bir organeldir. Spermatozoon’un ovum örtülerini ve zona pellucida’yı geçmesini sağlayan hyalurinidaz , corona penetrating enzim ve preakrozin akrozom tarafından salgılanır. Ovulasyondan sonra örtüleriyle birlikte fertilizasyon bölgesine gelen ovum’a yaklaşan kapasitasyonunu tamamlamış spermatozoon granulosa hücreleri ile temas sırasında akrozom reaksiyonuna uğrar. Bu reaksiyon sırasında akrozomun plazma memranı ile spermatozoon’un dış akrozomal membranı birçok noktada birbirleri ile birleşerek vesiküller oluştururlar. Bu veziküller arasında şekillenen boşluklardan akrozomal içerikteki enzimler dışarı çıkar. Akrozom reaksiyonu sonunda akrozom tümüyle parçalanır. Bu esnada spermatozoon’un başı sadece iç akrozomal memran ile sınırlanmış durumdadır. Akrozom reaksiyonu sonucu dışarı çıkan enzimler özellikle de hyalurinidaz granulosa hücrelerini birbirine bağlayan hyalurinik asit bağlarını eriterek örtülerin parçalanmasını ve spermatozoon’un kuyruk hareketleri ile bu örtüyü kolayca geçebilmesini sağlar.
Fertilizasyon
Ovum örtülerini geçen akrozom reaksiyonunu tamamlamış spermatozoon zona pellucida yüzeyine önce dik olarak yanaşır, sonra da başı boyunca yatay olarak uzanır. Bu uzanma sonucunda spermatozoon ile zona pellucida arasında bir bağlantı oluşur. Bu sırada perivitellin boşluğa I.kutup hücresi atılır. Zona pellucida’yı delen spermatozoon perivitellin boşluğu da çok kısa sürede geçerek vitellus’a yaklaşır ve baş kısmı vitellus yüzeyinde bulunan çok sayıda mikrovillus tarafından sarılır. Mikrovillus plazma membranları spermatozoon’un postakrozomal bölgesinin plazma membranı ile birleşirler. Plazma membranları arasındaki birleşme devam ederken kuyruk hareketleri ile spermatozoon’un tamamı perivitellin boşluğa girer.
Spermatozoon plazma membranı ile ovum mikrovillusları arasındaki birleşme ovum uyarımı denilen bir seri reaksiyona yol açar. Bu uyarım II.Meyoz bölünmenin metafaz safhasındaki ovumu uyararak bölünmesinin tamamlanmasına ve ikinci kutup hücresinin atılmasına neden olur. Spermatozoon plazma membranı ile ovum membranları arasındaki birleşme kademeli olarak kuyruğun posterior ucuna kadar sürer ve kuyruk dahil tüm spermatozoon ovum sitoplazmasına membransız olarak girmiş olur. Spermatozoon ovum sitoplazmasına girdiğinde nukleusunu çevreleyen membran parçalanır , kromatinler aktif hale geçer ve çevresini ovum sitoplazmasından köken alan yeni bir membran sararak erkek pronukleus’u şekillenir. Dişi kromatininin çevresini de ovum sitoplazması kökenli bir membran sararak dişi pronukleus’u şekillenir. Erkek ve dişi pronukleusları ovum sitoplazmasında merkeze doğru göçmeye başlarlar. Bu göç sırasında DNA duplikasyonu şekillenir. Pronukleuslar stoplazmanın merkezinde karşılaşırlar. Pronukleus mebranları erir, erkek ve dişiye ait kromozom grupları arasında birleşme şekillenerek fertilizasyon tamamlanır.
Döllenmiş Yumurtanın Taşınımı ve Nidasyon
Ovum döllendikten sonra 3-4 gün süreyle isthmus’ta kalır ve bölünür. Bu süre sonunda 8-16 hücreli hale gelen döllenmiş yumurta uterus-oviduct bağlantı yerini geçip cornu uteri’ye girer ve serbestçe dolaşarak bölünmeye devam eder. Bölünmenin 9.-10. günlerinde 60 ve daha üstü hücreye sahip olan blastosit aşamasındaki döllenmiş yumurta embriyo adını alarak zona pellucida’dan çıkar ve cornu duvarına yapışır. Bu sırada embrio varlığının sinyalini trofoblastin adlı bir madde salgılayarak anneye bildirir.
Bunun sonucunda uterus’tan prostaglandin salgısı durur ve yeni gelişen gebelik corpus luteum’undan (corpus luteum gravididatis) PGF2a salgılanır ve gebeliğin devamı sağlanır. Asıl plasentasyon gebeliğin 35.-40. günlerinde tamamlanır. Burada infertilite açısından erken embriyonik ölüm önem kazanır. Embriyo ölümlerinin %75 i gebeliğin 16. gününden önce, %25 i gebeliğin 16.-45. günleri arasında oluşur. O nedenle gebelik teşhisi en erken 45. günde yapılmalıdır. Ancak 45.günden sonra gebelik teşhis edilirse uygulanan suni tohumlama başarılı olmuş sayılır